
3D Ei
Ein Ei ist eine interessante biologische "Konstruktion". Insbesondere das Hühnerei gehört (bei uns) zu den wertvollen Grundnahrungsmitteln und ist in vielen Lebensmitteln enthalten. Hier sollen jedoch weder biologische noch ernährungs-physiologische Aspekte betrachtet werden.
Vielmehr soll die Frage beantwortet werden, wie man ein mathematisches 3D-Modell erzeugt, das dem Original recht nahe kommt.
Auch wenn es die Redewendung gibt "er/sie/es gleichen sich wie ein Ei dem anderen", so stellt man bei genauerer Betrachtung handelsüblicher Eier fest, dass es zusätzlich zur Größe (Gewicht), Farbe und Mustern (z.B. dunkle Flecken und Sprenkel bei braunen Eiern) auch bei der Form deutliche Unterschiede gibt; so sind z.B. manche Eier etwas "länglicher", andere hingegen "rundlicher".

Eigene Hühnerei-Modelle für den Unterricht
Für den Mathematikunterricht (Differential-/Integralrechnung) sollen geeignete Ei-Modelle gefunden werden. Allen Vogeleiern (s. Bild weiter unten) gemeinsam ist die Tatsache, dass der Eikörper rotationssymmetrisch um seine Längsachse ist. Es liegt daher nahe, das Ei als Rotationskörper zu modellieren. Im folgenden werden hierzu geeignete Randfunktionen mit möglichst wenigen Parametern aufgestellt, die bei der Rotation um die x-Achse das Ei entstehen lassen. Abschließend werden diese Modelle mit einem realen Ei verglichen.

Ei-Modell 1
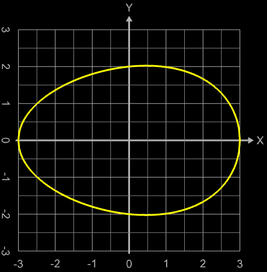
Da ein Ei im Prinzip ein Ellipsoid ist, jedoch mit unterschiedlicher Krümmung an den "Polen", wäre ein erster Ansatz der, als Randfunktion eine Ellipse zu verwenden, die jedoch entsprechend deformiert wird:

Mit a = 3 (halbe Höhe des Eies), b = 2 und c = 0.1 ergibt sich so der Querschnitt des Eies in der Abbildung.
Ei-Modell 2
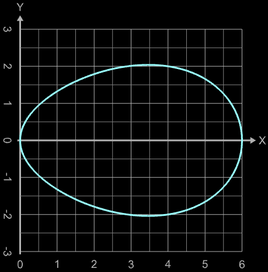
Für meinen Mathematikunterricht, bei dem auch die Volumenberechnung von Rotationskörpern ein Thema ist, habe ich folgende Randfunktion für die Rotation um die x-Achse aufgestellt:

Der Vorteil ist hier, dass das Integral bei der Volumenbestimmung
recht einfach zu berechnen ist, da die Wurzel wegfällt und somit nur ein einfaches Polynom zu integrieren ist (s. Volumen und Gewicht eines Hühnereies unter Rotationskörper/Volumen bei Rotation um x-Achse). Auch die Berechnung weiterer Größen, wie z.B. Integrationsgrenzen (ergeben die Länge des Eies) und maximaler Durchmesser stellen kein großes Problem dar.
Allerdings ist das entstehende Ei recht "spitz", wie z.B. mit a = 0.5, b= 0.2, c = 7.2 (s. Grafik).
Ei-Modell 3
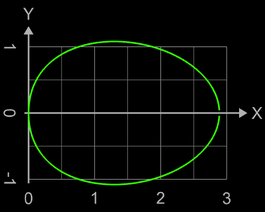
Hier wird im Vergleich zum vorigen Modell der Einfluss der 3. Potenz von x etwas "abgeschwächtt", wodurch u.a. die Spitze etwas runder wird:

Flexibler ist der folgende Ansatz:

Mit b = 1.5 ergibt sich die letzte Gleichung. Die Grafik zeigt die Randfunktion für b = 1.54 und c = 1.15.
Hier noch drei mit den obigen Modellen erzeugten Eierbeispiele:

Ei-Modell 4
Bei diesem Ei-Modell ist der Ausgangspunkt für die Randfunktion ein Kreis mit dem Radius 1, der durch

erzeugt wird. Der konstante Radius 1 wird durch eine Exponentialfunktion mit dem Parameter b ersetzt:

Mit b = 0 ergibt sich ein Kreis, während größere Werte für b diesen deformieren.
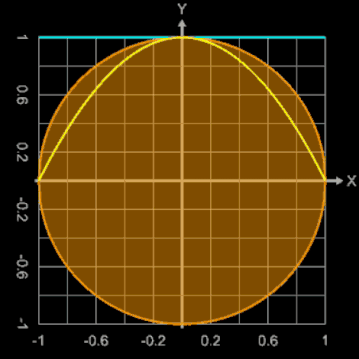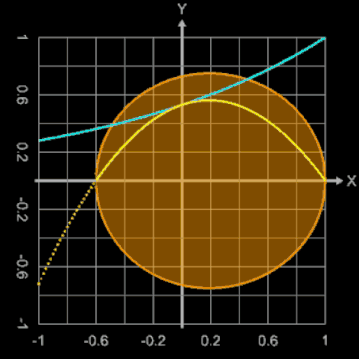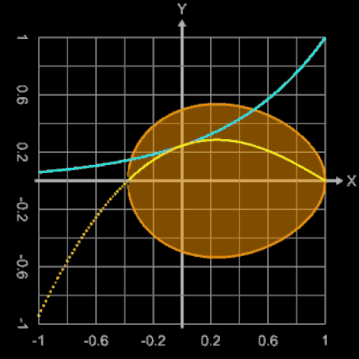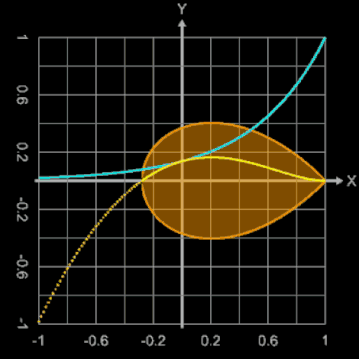
Die nebenstehende Animation zeigt dies für Werte von b zwischen 0 und 2. Dargestellt werden dabei

Brauchbare Werte für die Modellierung eines Hühnereies liegen für b im Bereich zwischen ca. 1.3 und 1.6. Zusätzlich kann mit dem Parameter a

der Graph der Randfunktion gestreckt / gestaucht werden. Das folgende Bild zeigt ein mögliches Ergebnis mit dem Ei-Modell 4, wobei a = 0.9 und b = 1.35 gewählt wurde.

Ei-Modell mit Goldenem Schnitt
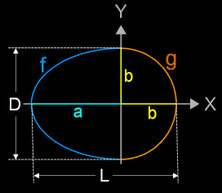
Das folgende Modell für ein Hühnerei besteht aus einer Halbkugel und einem halben Ellipsoid und beruht auf dem Goldenen Schnitt,. Es ist:
D = 2 b L = a + b a = Φ ∙ b
Φ = (1 + √5) / 2 ≈ 1.618
Somit gilt: (a + b) : a = a : b
Es kann als Rotationskörper erzeugt werden, indem die Funktionen f und g um 360° um die x-Achse rotiert werden mit

Es liegt nahe, die zuvor aufgeführten Modelleier mit real existierenden Eiern zu vergleichen und auch ein Gütekriterium für die Näherung aufzustellen. Dazu wäre folgendes Vorgehen erforderlich:
- Exaktes Fotografieren (d.h. mit perfekter Ausrichtung der Achsen und Bildebene) verschiedener Eier,
- Normierung der Bilder bezüglich der Größe,
- Ermitteln der Randpunkte eines Eibildes in einem Koordinatensystem
- Bestimmen der Parameter für das Näherungsmodell mit einem geeigneten statistischen Verfahren,
- Berechnen eines Gütekriteriums für die Abweichungen zwischen Ei und Modell.
All dies ist sehr aufwendig, so dass ich lediglich für zwei Eier aus meinem Kühlschrank die Modelle angewendet und dabei die Parameter durch Ausprobieren ermittelt habe:

Dies ist natürlich weder eine wissenschaftliche Herangehensweise noch eine praktikable Methode für das Bestimmen der Parameter. Zum anderen war der Gegenstand der obigen Betrachtungen lediglich ein Hühnerei.
Allgemein bei Vogeleiern gibt es jedoch eine große Vielfalt an Größen und Formen, von der archetypischen ovalen Form des Hühnereis über nahezu kugelförmige bis hin zu Eiern mit einem oder beiden spitzen Enden, von denen die folgende Abbildung eine Auswahl zeigt.

Quelle: www.shutterstock.com
Hauptformen von Vogeleiern (Nishiyama 2012)
Seit über einem Jahrhundert zerbrechen sich Biologen den Kopf, wie es bei der Bildung des Eies im Eileiter zu dieser Vielfalt kommt. Yutaka Nishiyama postulierte 2012, dass es vier verschiedene Hauptformen an Vogeleiern gibt [1]:
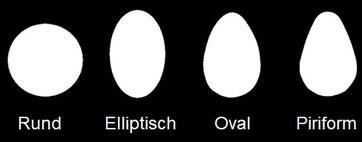
Die interessierte Leserschaft findet in dieser Publikation auch plausible Thesen dafür, warum die jeweilige Form existiert. So bauen beispielsweise diverse Seevögel, die an felsigen Steilküsten brüten, keine Nester, sondern legen vielmehr ihre Eier direkt auf den blanken Felsen. Durch die piriforme oder ovale Gestalt ihrer Eier können diese auf dem oft leicht abschüssigen Grund nicht wegrollen sondern bestenfalls kreiseln.
Bei den Modellen am Anfang dieser Seite war das Ziel, eine einfache Randfunktion y = f(x) aufzustellen, so dass der damit entstehende Rotationskörper einem (Hühner)ei sehr ähnlich sieht.
Im Folgenden sollen aber geeignete Funktionen gefunden werden, die real existierende Vogeleier beschreiben. Tatsächlich haben seit Mitte des zwanzigsten Jahrhundert einige Wissenschaftler sich damit beschäftigt, praktikable Modelle für Vogeleier zu entwickeln, wobei die Schwierigkeit darin bestand, dass beim Quantifizieren der Eiform keine eindeutige(n) Kenngröße(n) definiert war(en), die die gesamte Palette an Formen abdeckt.
Biggins et al. [17] fassen die Gründe, nach aussagekräftigen Darstellungen der Eiform zu suchen, wie folgt zusammen:
- Sie sind notwendig, um die Evolution und die adaptive Bedeutung verschiedener Formen zu verstehen.
- Die Existenz einer präzisen Formel für die Form stellt die Herausforderung dar, zu verstehen, wie der physikalische Prozess der Eibildung zu diesen interessanten, einfach zu beschreibenden Formen führt.
- Schließlich ist die Kenntnis der Form und Größe von Eiern für die kommerzielle Eierproduktion relevant.
Hühnerei-Modell von Romanoff (1949)
![Hühnerei-Modell von A.L. Romanoff & A. J.Romanoff [1]](https://image.jimcdn.com/app/cms/image/transf/dimension=338x1024:format=jpg/path/scee86bccd27a6ab2/image/i1264372e67286841/version/1773415055/image.jpg)
Im Jahr 1949 publizierten A.L. Romanoff und A.J. Romanoff ihr Werk "Das Vogel-Ei" [2], darin enthalten ein Modell für ein Standard- bzw. "ideales" Hühnerei (s. Grafik). Hierbei bedeuten:
Punkt M: Mittelpunkt der Ei-Länge L
Punkt O: Kreuzungspunkt
der maximalen
Länge L und maximalen Breite B
Strecke OP: halbe maximale Breite B/2
Punkt R: Punkt auf einem inneren Kreis
mit Mittelpunkt in O und Radius
OP (somit ist OP = OR)
Punkt S: der am
weitesten entfernte Punkt
am stumpfen Ende des Eies
Romanoff. 1949. The Avian Egg. New York, NY, USey & Sons Inc.
Hügelschäffer-Modell (1944)
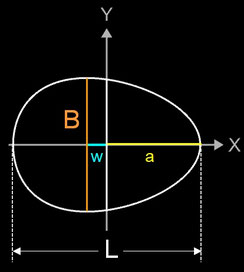
Ein vielversprechender Ansatz war das Modell von Hügelschäffer, das auf nur drei Parametern beruht. Der deutsche Ingenieur Fritz Hügelschäffer schlug als Erster eine eiförmige Kurve vor, die einem Ei ähnelt, indem er einen von zwei konzentrischen Kreisen entlang seiner x-Achse verschiebt und so eine asymmetrische Ellipse konstruiert [3] (bei [4] findet man dazu eine schöne Animation).
2 w x y² + b² x² + (a² + w²) y² - a² b² = 0

Hierin bezeichnen B = 2b die maximale Breite des Eies, L = 2a die Länge des Eies und w den Abstand zwischen zwei vertikalen Achsen, die der maximalen Breite und der halben Länge des Eies entsprechen.
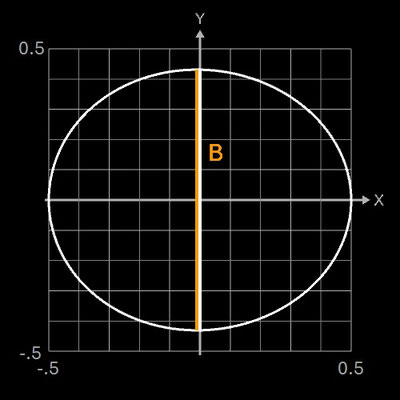
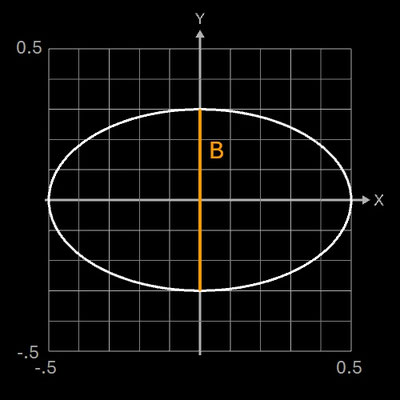
Falls im Hügelschäffer-Modell L = B ist, entspricht die Form einem Kreis, falls w = 0 ist, einer Ellipse. Daher eignet sich das Standardmodell von Hügelschäffer sehr gut für die drei klassischen Eiformen: kreisförmig, elliptisch und oval. Die folgenden Galerien zeigen je ein Beispiel-Ei [6] aus den vier Ei-Hauptformen und die mittels der Hügelschäffer-Methode berechneten Konturen der Eier. Die Abmessungen der Eier wurden so skaliert, dass alle die gleiche Länge von 1 haben.










Leider ist Hügelschäffers Modell zur Bestimmung der Konturen piriformer Eier nicht geeignet. So ist bereits visuell erkennbar, dass das berechnete Profil des Eies der Dickschnabellumme nicht dem realen entspricht. Narushin et al. (s. weiter unten auf der Seite) entwickelten das Modell weiter, so dass auch piriforme Eier modelliert werden können
Ursprünglich entwickelte Hügelschäffer sein Ei-Modell für die Luftfahrt. Auch heute noch wird dieses Modell für die Rumpfquerschnitte von Segelflugzeugen verwendet, wie in der folgenden Grafik dargestellt [7].

Weitere Anwendungen eiförmiger Kurven finden sich in der Architektur [9] sowie im Bauwesen, insbesondere in der Hydrologie. Abwassersysteme mit eiförmigen Rohrquerschnitten gewinnen aufgrund der Verfügbarkeit moderner Technologien in Städten zunehmend an Bedeutung [8].
Preston-Modell (1953, 1968, 1969)
F. W. Preston befasst sich in seiner 1953 erschienenen Publikation [10] mit der geometrischen Beschreibung von Vogeleiern und stellt fest, dass die herkömmliche Angabe von Länge und maximalem Durchmesser nicht ausreicht, um die Form eines Eies vollständig zu beschreiben, da die meisten Eier "an einem Ende dicker sind als am anderen". Er setzt sich das Ziel, alle denkbaren Eiformen mittels einer mathematischen Gleichung präzise abzubilden. Hierzu nutzt Preston zur Darstellung von Eiprofilen parametrische Gleichungen und entwickelt sein Modell schrittweise:
- Mit den Parametern Länge L und maximale Breite B lässt sich eine Ellipse beschreiben:
x (Φ) = L∙sin(Φ), y(Φ) = B∙cos(Φ)
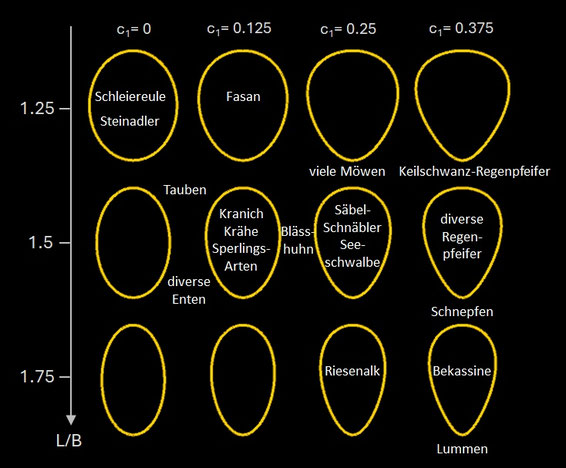
- Mit einem weiteren Parameter c1 kann die einfache Eiform ("simple ovoid") mit ihrer Asymmetrie zwischen dem stumpfen und dem spitzen Ende dargestellt werden:
x (Φ) = L∙sin(Φ), y(Φ) = B∙cos(Φ) ( 1+c1∙sin(Φ) )
Die folgende Grafik zeigt die so erzeugten Eiformen und zugehörigen Vogelarten.
- Für die meisten Eier ("standard avian") ist ein weiterer Parameter c2 nötig, der das Ei "bikonisch" macht, d.h. beide Enden wirken etwas kegelförmiger:
x (Φ) = L∙sin(Φ), y(Φ) = B∙cos(Φ) ( 1+c1∙sin(Φ) + c2∙sin²(Φ) )
- Piriforme (birnenförmige) Eier ("alcid ovoid"), wie z.B. die der Trottellumme oder des Kiebitzes, benötigen noch eine weitere Konstante c3, um die starke Zuspitzung an einem Ende korrekt zu erfassen:
x (Φ) = L∙sin(Φ), y(Φ) = B∙cos(Φ) ( 1+c1∙sin(Φ) + c2∙sin²(Φ) + c3∙sin³(Φ) )
Die etwas "unhandliche" parametrische Gleichung lässt sich in eine kartesische Form bringen, um z.B. das Volumen oder die Oberfläche des Eies als Rotationskörper zu berechnen:

Normiert man die Gleichung mit L = 1, ergibt sich:

Die folgende Grafik zeigt den Einfluss der Parameter c0 bis c3 auf die Eiform [12a].
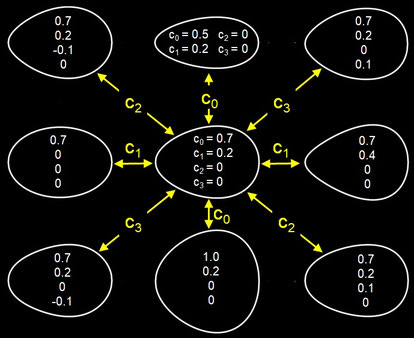
Um die notwendigen Werte für diese Parameter zu gewinnen, entwickelte Preston eine spezielle Profil-Kopiermaschine. Er unterteilte die Länge L eines Eies in
acht gleichbreite Bereiche und ermittelte mit seiner Maschine die sieben Durchmesser D1 bis D7 entlang der Längsachse (s. folgende linke Grafik), um die Werte für c0 bis c3
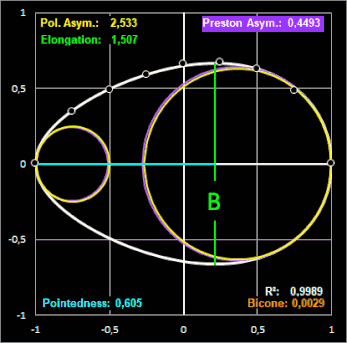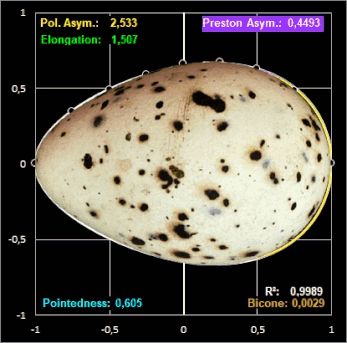
mittels der Methode der kleinsten Fehlerquadrate zu berechnen. Für das Beispiel des Eies der Königsseeschwalbe (Thalasseus maximus) ergeben sich die
Parameter zu c0 = 0.64643317,
c1 = 0.23686193, c2 = c3 = 0 bei einem Bestimmtheitsmaß von R² = 0.998913 und mit dem grafischen Ergebnis in der folgenden rechten
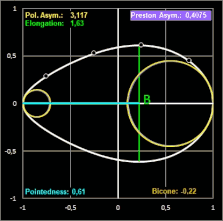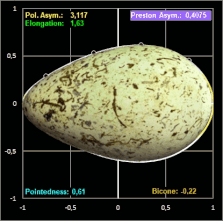
Grafik. Die Definition der Parameter Polare Asymmetry und Pointedness finden Sie weiter unten bei Biggin et al. Studie
(2018).

Seine Studie zeigt, dass selbst "extrem" geformte Eier mit nur wenigen mathematischen Termen sehr genau (auf 0,1 bis 0,2 mm präzise) dargestellt werden können. Hier zwei Beispiele mit je einem Ei des Halsbandfrankolin (Frankolinus frankolinus) und der Trottellumme (Uria aalge).
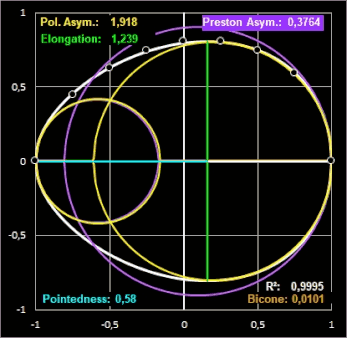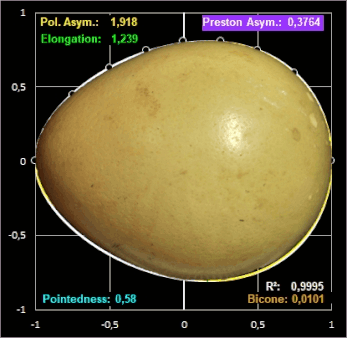

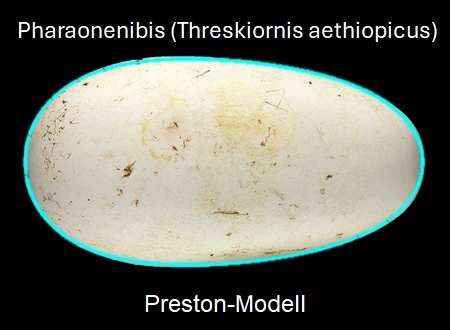
Das linke Bild in der folgenden Galerie zeigt das ungewöhnlich geformte Ei des Pharaonenibis (Threskiornis aethiopicus) und die mit dem Preston-Modell erzeugte Kontur (cyan). Für das gleiche Ei liefern das Modell von Hügelschäffer und das Universalmodell von Narushin et al. vergleichsweise schlechtere Ergebnisse:


In einer späteren Publikation von 1968 [11] führt Preston zur Beschreibung der Form eines Eies die drei voneinander unabhängigen, dimensionslosen Formindices Elongation (Streckung), Asymmetry (Asymmetrie) und Bicone (Bikonizität) ein. Dazu werden neben der herkömmlichen Länge L und maximalen Breite B die Krümmungsradien des stumpfen ρB und des spitzen Endes ρP mit Hilfe eines Sphärometers [12b] gemessen und wie folgt in Beziehung gesetzt:
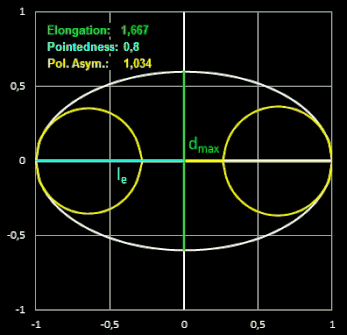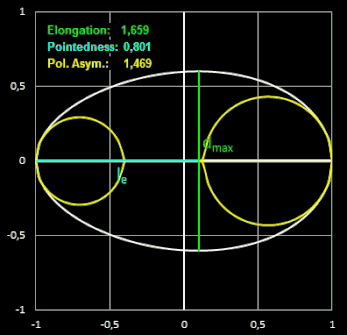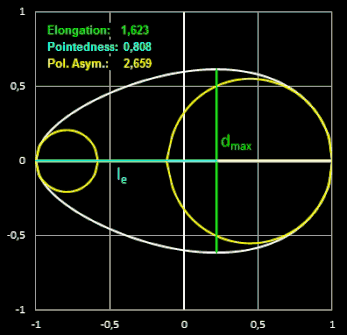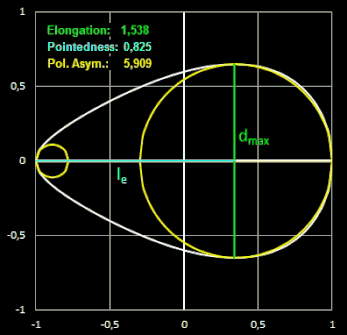
- Elongation = L / B
- Asymmetry = (ρB - ρP) L / B²
- Bicone = (ρB + ρP) L / B² - 1
Die Elongation beschreibt, wie stark ein Ei in Richtung der Längsachse gestreckt ist. Bei einem Wert nahe 1 hat das Ei fast die Form einer Kugel.
Anschaulich beschreibt die Asymmetry, wie stark ein Ende des Eies größer oder dicker ist als das andere. Bei geringer Asymmetrie ist das Ei an beiden Enden fast gleich geformt, bei einer hohen Asymmetrie ist ein Ende deutlich stumpfer (breiter), während das andere Ende spitz zuläuft (diese Eiform wird als piriform oder birnenförmig bezeichnet). Ist die Asymmetry gleich Null, so lässt sich zunächst nichts über die Form der Ei-Enden aussagen. Diese können sehr unterschiedlich sein, wie dies die folgenden zwei Beipiele fiktiver Ei-Modelle zeigen (die gestrichelten Linien sind die Konturen der Ellipse mit den Halbachsen B/2 und L/2).
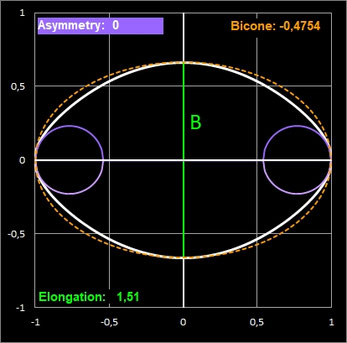
Elongation (Dehnung) ist das Verhältnis der Länge L zum maximalen Durchmesser B.

Der Formindex Bicone schließlich beschreibt anschaulich, inwieweit beide Enden eines Eies spitzer oder stumpfer sind, als man es bei einem normalen Ellipsoid mit gleicher Länge L und Breite B erwarten würde. Die meisten Vogeleier tendieren dazu, etwas spitzer zulaufend zu sein als ein Ellipsoid. Dies wird mathematisch mit einem negativen Vorzeichen versehen, da das Ei dadurch weniger Volumen hat als die Vergleichsform. Bei einigen Familien wie Kolibris oder Albatrossen sind beide Enden "stumpfer" bzw. bauchiger. Diese Eier haben mehr Volumen als ein idealisiertes Ellipsoid.
Ein Wert von Null bei Asymmetry und Bicone entspricht einem perfekten Ellipsoid.
Hier noch drei Beispiele mit dem Ei [6] des Löffelreihers (Bicone negativ), des Sterntauchers (Bicone fast 0) und der Sternelfe, einer Kolibri-Art (Bicone positiv):

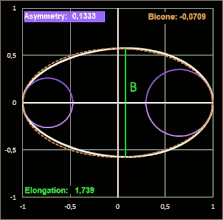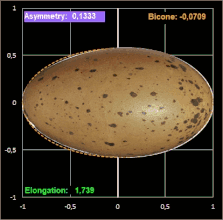
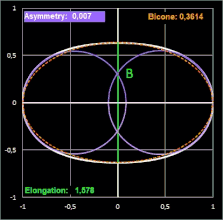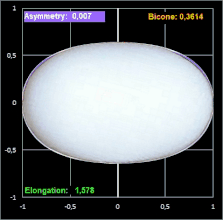
Prestons Studie zeigt, dass die Einbeziehung der Endkrümmungen eine nahezu vollständige Definition der Größe und Form eines Eies ermöglicht. Somit liefert seine Arbeit die theoretische und praktische Grundlage für die systematische Erfassung von Eiformen in ornithologischen Handbüchern.
Biggins et al. Studie (2018)
In der interessanten Publikation [13]
geben die Verfasser zunächst einen ausführlichen und kritisch-historischen Rückblick über Methoden zur mathematischen Beschreibung der vielfältigen Formen von Vogeleiern. Sie greifen dann das Modell von Preston (s.o.) auf, bei dem
das Eiprofil
y (x) als Modifikation eines Kreises durch ein Polynom 3. Grades dargestellt wird:

Da die rein mathematischen Preston-Parameter c0, ..., c3 schwer intuitiv zu verstehen sind, leiten die Autoren daraus drei neue, biologisch interpretierbare, charakteristische Formindices für Vogeleier ab:
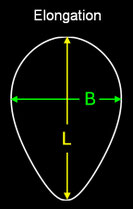
Elongation
("Ausdehnung") ist das Verhältnis der Länge L des Eies zu dessen maximaler Breite B.

Pointedness ("Zuspitzung") ist das Verhältnis der Länge LP
vom maximalen Durch-messer bis zum weiter entfernten Ende, zu der Gesamtlänge L.

Polar Asymmetry ("Polare Assymetrie") ist das Verhältnis des Durchmessers
DB des
größten
Kreises, der in den Eiquerschnitt passt und dabei das Ei an seinem stumpfen Pol berührt, zum Durchmesser des größten Kreises DP innerhalb
des Eiquerschnitts, der den spitzeren Pol berührt.
Größere Werte dieser Formindices entsprechen größeren Abweichungen von einer Kreisform. In der folgenden Grafik [13] mit unterschiedlichen Eiformen kann man den Zusammenhang zwischen den Formindices und der Eiform gut erkennen (alle Eiformen wurden auf die gleiche Länge skaliert). Zugrunde lagen die Messungen an 132 Eiern von 10 Vogelarten.

1 - Braunliest (Halcyon smyrnensis)
2 - Adeliepinguin (Pygoscelis adeliae)
3 - Krauskopfpelikan (Pelecanus crispus)
4 - Rosaflamingo (Phoenicopterus roseus)
5 - Südlicher Streifenkiwi (Apteryx australis)
6 - Zwergtaucher (Tachybaptus ruficollis)
7 - Königsseeschwalbe (Thalasseus maximus)
8 - Königspinguin (Aptenodytes patagonicus)
9 - Fasanblatthühnchen (Hydrophasianus chirurgus)
10 - Trottellumme (Uria aalge)
Zwar hatte Preston bereits den Formindex Asymmetrie definiert [11]; der Formindex Polar Asymmetry von Biggins et al. wird hier jedoch direkter als das Verhältnis der beiden Krümmungsradien definiert und beschreibt somit, wie viel "stumpfer" das dicke Ende im Vergleich zum spitzen Ende des Eies ist. Der Index ist eine biologisch intuitivere Weiterentwicklung basierend auf der mathematischen Exzellenz von Prestons ursprünglicher Arbeit.
In den folgenden beiden Animationen werden für verschiedene Eiformen die oben definierten Formindices berechnet:


Intermezzo
Etwas irritiert bin ich über den Formindex Polare Asymmetrie. Diesen definieren Biggins et al. "als das Verhältnis der beiden größten Kreise, die in die Ei-Kontur passen und dabei das stumpfe bzw. spitzere Ende des Eies berühren".
Um die beiden Radien rB der In-Kreise am stumpfen Ende und rP am spitzeren Ende zu bestimmen, habe ich in EXCEL eine Näherung programmiert, bei der jeweils der Radius eines Kreises mit Berührpunkt an einem der Enden sukzessive in kleinen Schritten erhöht wird, solange für einen Schritt alle y-Werte des Kreises kleiner als die der Ei-Kontur sind. Diese beiden Radien nennen Biggins et al. RB und RP (s. [17], Tabelle 1).
In dieser Tabelle heißen die beiden Radien bei den Forneln von Preston gleich, obwohl diese die Krümmungsradien an den Ei-Enden darstellen, die sie lt. [11] per Sphärometer ermitteln.
Bestimmt man den Krümmungsradius r ausgehend von Prestons Gleichung (6) in [10], so ergibt sich für diesen:

Ich habe für einige Eiformen jeweils beide Varianten an "Krümmungskreisen" ermittelt und festgestellt, dass sie nahezu gleich sind. Für größere Werte von RB / RP und rB / rP (im Beispiel unten ab ca. 3) wird der berechnete Krümmungsradius rB jedoch größer als RB. Somit wird auch der Index Asymmetrie (rB / rP) größer als der Index Polare Asymmetrie (RB / RP), wie dies beispielhaft folgende Animation zeigt.
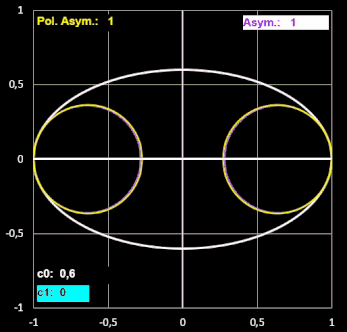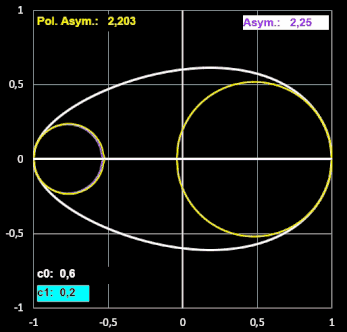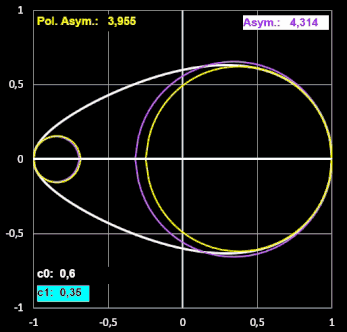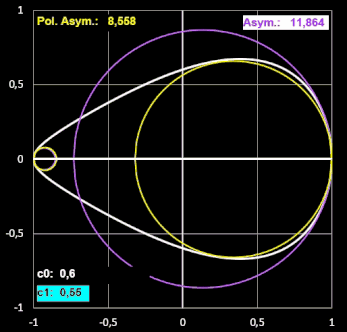
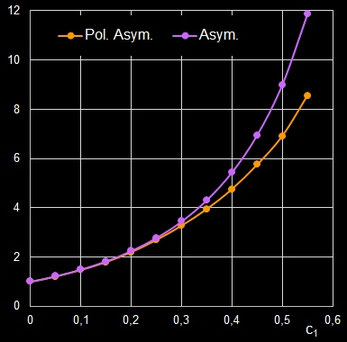
Ein zentraler Beitrag der Arbeiten von Biggins et al. ist die Bereitstellung eines R-Programms, das für Ei-Fotographien automatisch die Umrisse (Silhouetten) des Eies extrahiert und die Preston-Parameter durch Minimierung der Fehlerquadratsumme (Least Squares Fit) berechnet. Durch das neue Tool können Forscher nun große Mengen an digitalen Bildern effizient verarbeiten und Formmerkmale exakt quantifizieren, was für evolutionäre und ökologische Studien von großer Bedeutung ist.
Einige digitalisierte Eibilder, die Herleitung der Modellparameter, einen statistischen Vergleich mit dem Original sowie die verwendeten Programme (Programmiersprache "R") wurden in [14] publiziert.
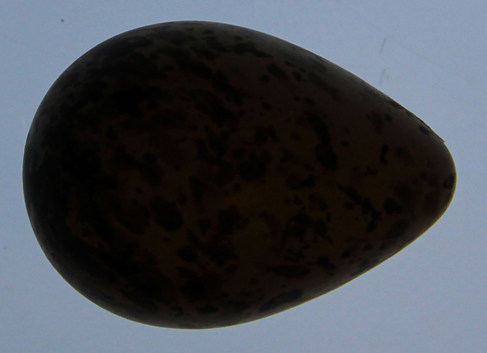
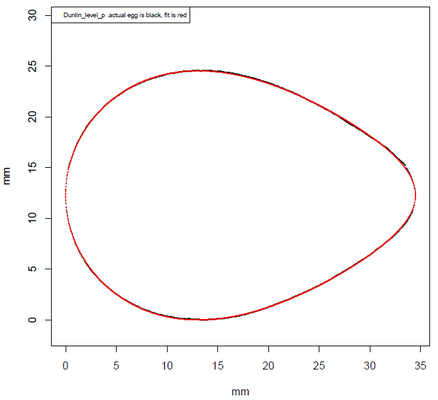
Ich habe aus diesen Daten das Ei des Alpenstrandläufers (Calidris alpina)
herausgepickt. Für x-Werte im Bereich [-1, 1] lauten die Parameter: c0 = 0.68278…, c1 = 0.22139…, c2 = -0.05116…, c3 = 0.03457…
Die folgende Galerie zeigt von links nach rechts: Alpenstrandläufer (www.wikipedia.de), sein Ei (www.wikipedia.de), Eifoto zur Erfassung des Randes ([14]), grafischer Vergleich Rand des Eies mit Modell ([14]) und das von mir mit den obigen Parametern berechnete Ei, versehen mit einem zufälligen Farbmuster (vgl. 3D Texturen unter Meeresschnecken und Muscheln / Modell B. Frassek).



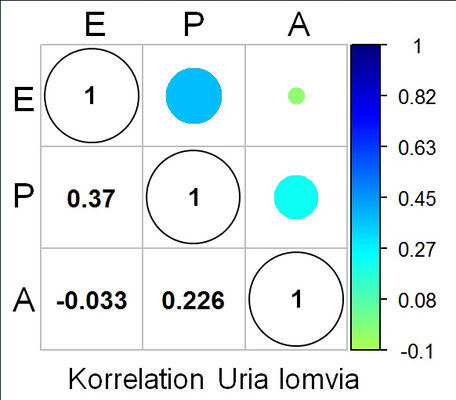
Insbesondere für die drei Seevogelarten Trottellumme, Dickschnabellumme und Tordalk, die an steilwandigen Küstenformationen brüten, hat J.D. Biggins für deren piriforme Eier (s. folgende Galerie) die drei charakteristischen Kenngrößen (s.o.) ermittelt und deren Korrelation berechnet [14, Supplementary Material].



Fotos von Klaus Rassinger und Gerhard Cammerer, Museum Wiesbaden
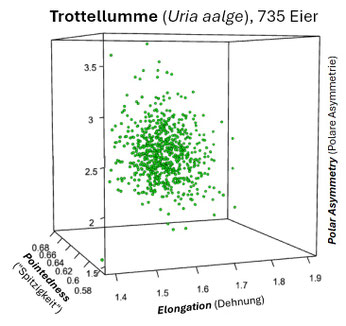
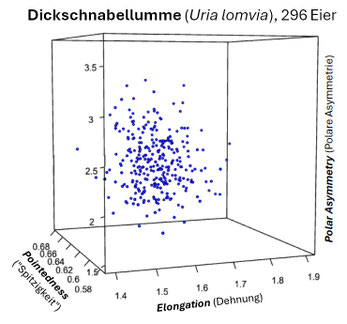

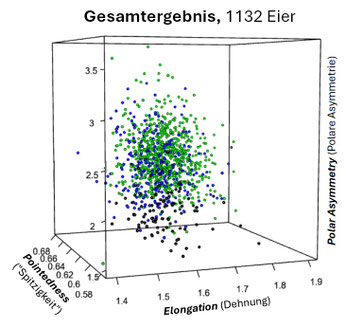
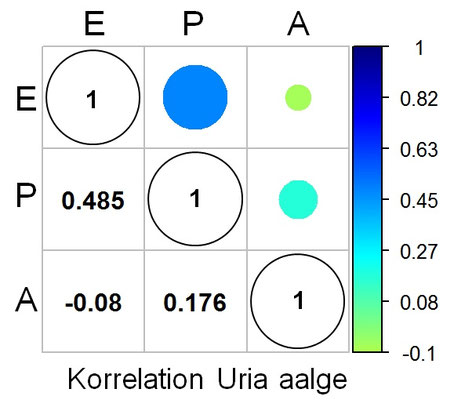
Die folgenden drei Grafiken zeigen für diese Eier die Shearman-Korrelation für die drei Kenngrößen E: Elongation (Ausdehnung), P: Pointedness (Spitzigkeit) und A: Polar Asymmetry (Polare Asymmetrie). Insgesamt ist die Korrelation aber eher sehr schwach.

Narushin et al.-Modell (2021)

Narushin et al. greifen das Modell von Hügelschäffer (s.o.) auf und entwickeln es dahingehend weiter, dass auch piriforme Eier modelliert werden können [15]. Hierzu führen sie einen weiteren Parameter DL/4 ein (Durchmesser des Eies bei ¼ seiner Länge vom spitzen Ende aus gemessen, s. Grafik rechts) und leiten folgende Funktion zur Darstellung der Kontur eines piriformen Eies her:
 Die
folgenden Galerien zeigen einige Beispiele für Konturen piriformer Eier [6],
die ich mit diesem Modell berechnet habe.
Die
folgenden Galerien zeigen einige Beispiele für Konturen piriformer Eier [6],
die ich mit diesem Modell berechnet habe.




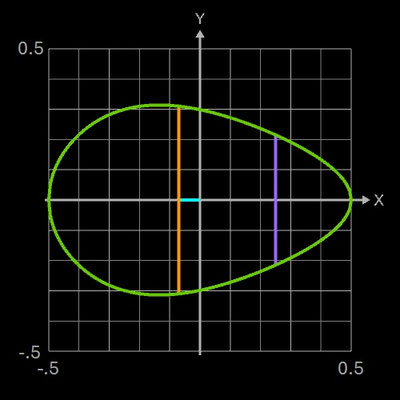







In der obigen rechten Grafik erkennt man deutlich den Unterschied der mit den Funktionen fH
(Hügelschäffer, weiß) und fP (piriformes
Ei-Modell, grün) berechneten Konturen. Um auch "Zwischenformen" von Eiern modellieren zu können, leiten Narushin et al. folgende Funktion her (die zugehörigen Supplements ihrer Publikation mit
den aufwendigen Berechnungen konnte ich downloaden und stelle sie für die interessierte Leserschaft als Download bereit [16]):

Die folgenden Galerien zeigen einige Beispiele für Eier [6], deren Konturen ich mit dieser Funktion berechnet habe.







Biggins et al. Studie (2022)
Preston stellte in seiner Publikation von 1953 [10] fest, dass bereits vier gemessene Eidurchmesser zusammen mit der Eilänge L ausreichten, um die vier Parameter c0 bis c3 in seiner Gleichung

zu schätzen, leitete jedoch keine explizite Formel darauf basierend ab.

In ihrer Publikation [17] und Supplement [18] von 2022 unterteilen Biggins et al. die Länge L des Eies in Achtel (Oktile): D1 ist der Eidurchmesser im ersten Oktil, D2 im dritten, D3 im fünften und D4 im siebten (s. Grafik).
Diese vier zu messenden Durchmesser werden als Bruchteile der Eilänge durch di = Di / L ausgedrückt. Für diese vier Werte leiten Biggins et al. auf Basis eines allgemeinen linearen Modells und multiplen Regression die folgende Gleichung für die Eiform her, die Prestons obiger Gleichung mit Parameterschätzungen aus vier spezifischen Messpunkten entspricht, und die sie "Four-Diameter Equation" (Vier-Durchmesser-Gleichung) nannten:

Die Formel liefert den Radius y des Eies an der Stelle x mit −1 ≤ x ≤ 1, wobei die Enden des Eies bei x = −1 und x = 1 liegen (d.h. für die Eilänge gilt L=2).
Soll der Kenngröße Polare Asymmetrie mittels der Krümmungsradien rB und rP bestimmt werden (s. dazu auch oben bei Biggins et al. Studie 2018), ergeben sich diese gemäß:

mit

Für einige Vogeleier (Fotos [6]) mit recht unterschiedlichen Formen habe ich deren Konturen mit der Vier-Durchmesser-Gleichung (im Folgenden 4-Diam-Equ genannt) bestimmt und war verblüfft wegen der recht hohen Genauigkeit des Modells.
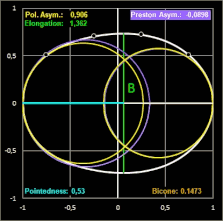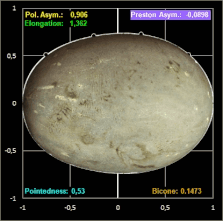

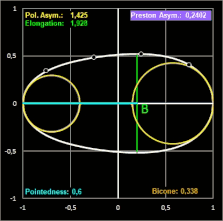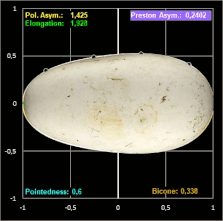


Das Hauptanliegen ihrer Publikation war es für Biggins et al. jedoch, in einem umfassenden Vergleich verschiedener Studien zur Modellierung von Vogeleiern zu zeigen, dass alle seit den 1950er Jahren veröffentlichten Methoden zur Schätzung der Eiform in unterschiedlichem Maße ungenauer als Prestons ursprüngliche Formulierung sind.

Biggins et al. untersuchten, wie genau verschiedene Modelle die Konturen eines Eies widergeben.
Für ein breites taxonomisches Spektrum verwendeten sie Fotografien von jeweils einem Ei von 25 Arten mit piriformen und 25 Arten mit nicht-piriformen Eiern; diese glichen sie mit den Daten von Montgomerie et al. (5380 Eier von 956 Arten) [19] ab. Die ausgewählten Arten repräsentieren eine große Bandbreite an Eiformen [18], was für einen überzeugenden Methodenvergleich notwendig ist.
Die Ergebnisse stellten sie in nebenstehender Übersichtsgrafik zusammen. Dort zeigen Boxplots den Fehler als Prozentsatz der Eilänge. Die Boxen umschließen oberes und unteres Quartil, die dicke Linie innerhalb stellt den Meridian dar, die Whisker reichen bis zum Extremwert innerhalb des 1,5-fachen Interquartilsabstands und extremere Werte sind einzeln markiert.
Die Anzahl dimensionsloser Parameter, die in einem Modell zur Spezifizierung der Form benötigt werden, ist in rot dargestellt.
Die Modelle sind anhand des Gesamtmedians pro Modell geordnet: vom ungenauesten (oben) zum genauesten (unten).
Für piriforme Eier liefert Prestons Gleichung [P6] (s.o.) mit Schätzungen, die aus dem vollständigen Eiprofil abgeleitet wurden, bessere Ergebnisse als alle anderen Methoden! Die nächstliegende Alternative ist die oben eingeführte Vier-Durchmesser-Gleichung.
Für nicht-piriforme Eier sind die Vier-Durchmesser-Gleichung sowie zwei andere nahezu so effektiv wie Prestons Gleichung [P6]. Die relativ schwache Leistung der Eiformformel in Narushin et al. 2021 [15] ist bemerkenswert, denn obwohl die Verfasser Preston (1953) zitieren, behaupten sie, die gesuchte universelle Formel für die Eiform entdeckt zu haben. Jedoch ist sowohl für piriforme als auch für nicht-piriforme Eier Prestons Gleichung [P6] allen anderen Methoden signifikant überlegen.
Eventuell erwiesen sich seine Gleichungen für Ornithologen als schwer verständlich, letztlich wurden sie weitgehend ignoriert. Trotz der Effektivität und Genauigkeit von Prestons ursprünglichen Vorschlägen von 1953 erschienen in den darauf folgenden 70 Jahren zahlreiche Veröffentlichungen mit anderen mathematischen Beschreibungen. Laut Biggins et al. beziehen sich diese Publikationen zwar häufig auf Prestons Arbeit, überprüfen aber weder die Genauigkeit ihrer Formulierungen hinsichtlich der Darstellung realer Eierformen über die gesamte Bandbreite interspezifischer Variationen hinweg, noch vergleichen sie die Effektivität ihrer Vorschläge mit Prestons Ansatz. Oftmals wurde viel Aufwand in die "Lösung" eines Problems investiert, das Preston bereits 1953 umfassender und effektiver gelöst hatte.
Da alle seit den 1950er Jahren veröffentlichten Methoden zur Schätzung der Eiform in unterschiedlichem Maße ungenauer als Prestons ursprüngliche Formulierung sind, postulieren Biggins et al., dass Prestons Ansatz zur Eiformbestimmung in zukünftigen Studien als Standardmethode verwendet werden sollte und neue Vorschläge für Formeln zur Bestimmung der Eiform nur dann in Betracht gezogen werden sollten, wenn sie mindestens so gut wie Prestons universelle Formel funktionieren und zusätzliche biologische Erkenntnisse oder andere spezifische Vorteile bieten.
Zum Abschluss dieser Seite über Modelle von Vogeleiern hier noch ein paar Eier unterschiedlicher Größen und Formen, die allesamt Seevögeln entstammen [19]:
Von links nach rechts: Madeirawellenläufer (Hydrobates castro), Bulwersturmvogel (Bulweria bulwerii), Schuppensturmtaucher (Puffinus lherminieri), Keilschwanz-Sturmtaucher (Ardenna pacifica), Gelbnasenalbatros (Thalassarche chlororhynchos), Kurzschwanz-Sturmtaucher (Ardenna tenuirostris),
Weißkinn-Sturmvogel (Procellaria aequinoctialis), Riesensturmvogel (Macronectes giganteus), Dunkler Sturmtaucher (Ardenna grisea), Wanderalbatros (Diomedea exulans)
Quellenverweise
[1] Nishiyama, Y. (2012) The mathematics of egg shape, Int. J. Pure Appl. Math. 78: 679–689
[2] Romanoff, A.L., Romanoff A.J. (1949) The Avian Egg, New York, NY, USA: John Wiley & Sons Inc.
[3] F. Hügelschäffer (1944) Das Ei als mathematisches Problem, VDI-Zeitschrift, Band 88, Nummer 11/12
[4] https://www.mathcurve.com/courbes2d.gb/oeuf/oeuf.shtml
[6] https://commons.wikimedia.org/wiki/Category:Eggs_of_the_Natural_History_Collections_of_the_
Museum_Wiesbaden
[8] M. Petrovic, B. Malesevic (2023) Hügelschäffer Egg Curve And Surface, Applicable Analysis and
Discrete Mathematics 17(1):179-196, https://doi.org/10.2298/AADM220526027P
[10] Preston, F. W. (1953) The Shapes of Birds' Eggs, The Auk: Vol. 70: Iss. 2, Article 5.
[11] Preston, F. W. (1968)
The Shapes of Birds' Eggs: Mathematical Aspects,
The
Auk: Vol. 85: Iss. 3, Article 10
[12] Preston, F. W. (1969)
Shapes of Birds' Eggs: Extant North American Families,
The
Auk: Vol. 86: Iss. 2, Article 5
[12a] Z. Barta, T. Székely (1997) The optimal shape of avian eggs, Functional Ecology 1997 11, 656-662
[13] Biggins J.D., Thompson J.E., Birkhead T.R. (2018) Accurately quantifying the shape
of birds’ eggs,
Ecol Evol. 2018; 8: 9728–9738, https://doi.org/10.1002/ece3.4412
[14] https://datadryad.org/resource/doi:10.5061/dryad.8kv2b20
[15] Narushin V. G., Romanov M. N., Griffin D. K. (2021) Egg and math: introducing a universal formula for
egg shape, Ann. N.Y. Acad. Sci. 1505 (2021) 169–177, DOI:10.1111/nyas.14680
[16] Narushin V. G., Romanov M. N., Griffin D. K. Supplements S1-S4, ZIP-Datei
[17] Biggins
J.D., Montgomerie R., Thompson J.E., Birkhead T.R. (2022) Preston’s universal formula for
avian egg shape, Ornithology, Volume 139, pp. 1–8,
https://doi.org/10.1093/ornithology/ukac028
[18] Biggins et al. (2022) Supplements (Daten und Analysen), 443 MB (!),
https://doi.org/10.5061/dryad.547d7wmbz
[19] Montgomerie, R., Hemmings, N., Thompson, J.E. et al. (2021) The shapes of birds’ eggs:
evolutionary constraints and
adaptations, The American Naturalist, 198 (6). E215-E231,
ISSN: 0003-0147, https://doi.org/10.1086/716928